
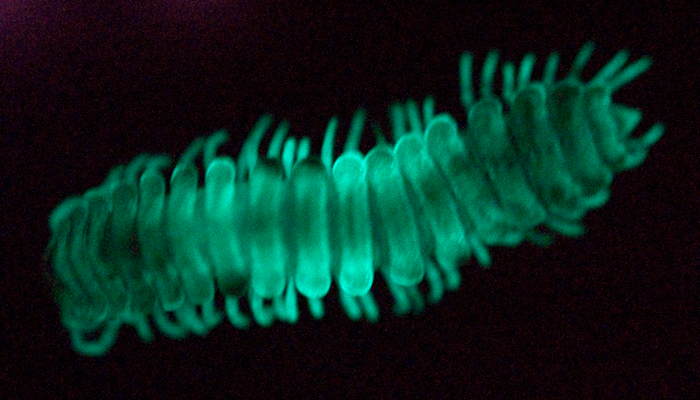
In the Sierra Nevada Mountains of California, bioluminescent millipedes scatter upon the forest floor and on a moonless night, “resemble a starry sky” (Tiemann, 1969). The most remarkable feature of these millipedes is their ability to glow at a wavelength of 495 nm. This is only one of two known instances of bioluminescence in the entire millipede class Diplopoda. Bioluminescence in millipedes is restricted to only eight species of the genus Motyxia, which are endemic to a very small area in the Sierra Nevada Mountains, and Paraspirobolus lucifugus (a distantly related species in the order Spirobolida) from Japan, Taiwan and the Federated States of Micronesia.
The function of luminescence in Motyxia remained uncertain. Authors speculated that the role of the emitted light may be a nocturnal warning signal to announce the presence of a cyanide-based chemical defense, as reflected color does in closely related millipedes in Appalachia. Still, others suggested that the luminescence serves no adaptive function.
Our current research program aims to discover the evolutionary circumstances under which this unique adaptive innovation arose. The first part of the project consists of molecular phylogenetics as a foundation to address monophyly of the genus, and as a basis for new species descriptions. Did luminescence evolve multiple times, and under what circumstances? The second part of the project is a field test of luminescence and whether it functions as a nocturnal warning signal. Currently, we are using next generation transcriptome sequencing to understand the DNA-level differences between luminescent and closely related non-luminescent taxa.
[Above, the brightest species Motyxia sequoiae photographed in its own light (174 s exposure) from Giant Sequoia National Monument, California.]
Molecular phylogenetics of Motyxia and relatives (Xystodesmidae)
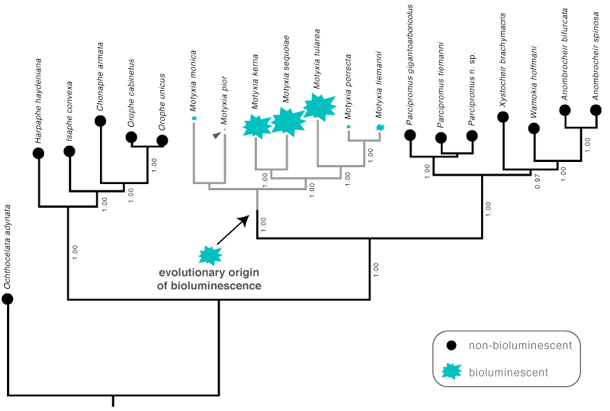
^ Phylogeny of Motyxia species and close relatives. Bioluminescence has a single evolutionary origin in the millipede order Polydesmida (arrow).
In this figure, starbursts indicate presence and relative intensity of bioluminescence, circles denote absence of bioluminescence. Ancestral origin of bioluminescence is indicated by a solid arrow at the base of the Motyxia clade. Weak bioluminescent intensity of Motyxia pior emphasized with a triangle. Motyxia sequoiae display the brightest bioluminescence of the genus. Phylogeny estimated with every known species of Motyxia (spare one) and representatives from five closely related tribes.
Testing bioluminescent aposematism in millipedes
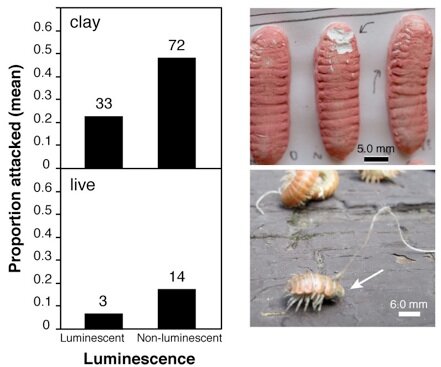
^ Results of a field experiment to test the prediction that luminescent millipedes are attacked less often than non-luminescent millipedes.
In this figure, the bar graphs show the experiment’s results: the proportion of millipedes attacked versus luminescence. Non-luminescent millipedes were attacked more often than luminescent millipedes. Images show rodent incisor marks in clay millipedes, and live millipede (arrow) with anterior segments 1–14 missing after predator attack.
Figures adapted from: Marek et al. (2011) Bioluminescent aposematism in millipedes. Current Biology, 21, R680-R681. [Open access]
This research is supported by a National Science Foundation Phylogenetic Systematics grant to Paul Marek (DEB-1119179)

{kind=link}